Principles of Metabolic
Regulation
chapter
15
S-173
1. Measurement of Intracellular Metabolite Concentrations Measuring the concentrations of
metabolic intermediates in a living cell presents great experimental difficulties—usually a cell must
be destroyed before metabolite concentrations can be measured. Yet enzymes catalyze metabolic
interconversions very rapidly, so a common problem associated with these types of measurements is
that the findings reflect not the physiological concentrations of metabolites but the equilibrium con-
centrations. A reliable experimental technique requires all enzyme-catalyzed reactions to be instanta-
neously stopped in the intact tissue so that the metabolic intermediates do not undergo change. This
objective is accomplished by rapidly compressing the tissue between large aluminum plates cooled
with liquid nitrogen (190 C), a process called freeze-clamping. After freezing, which stops enzyme
action instantly, the tissue is powdered and the enzymes are inactivated by precipitation with perchlo-
ric acid. The precipitate is removed by centrifugation, and the clear supernatant extract is analyzed
for metabolites. To calculate intracellular concentrations, the intracellular volume is determined from
the total water content of the tissue and a measurement of the extracellular volume.
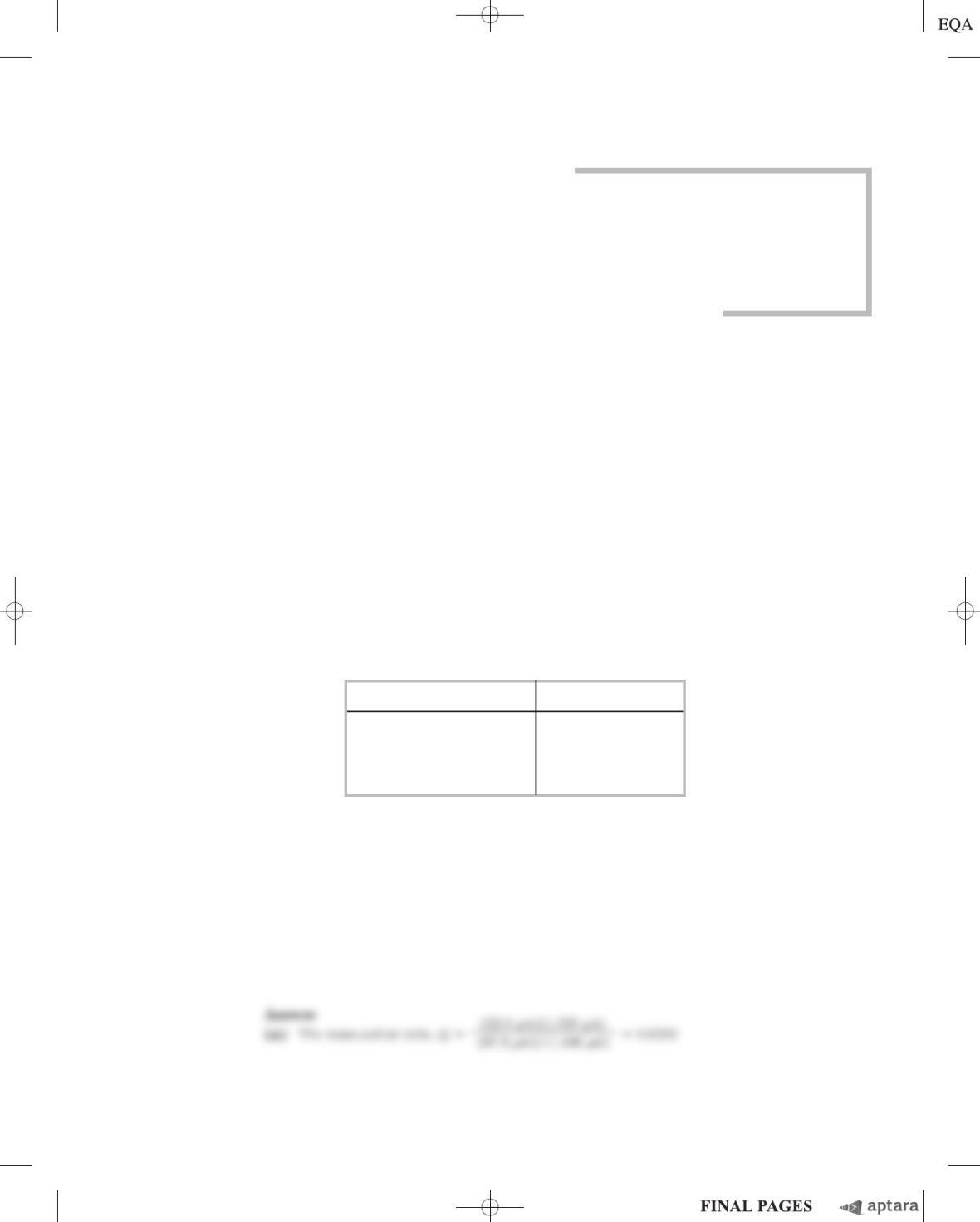
The intracellular concentrations of the substrates and products of the phosphofructokinase-1 reac-
tion in isolated rat heart tissue are given in the table below.
Metabolite Concentration m
M
*
Fructose 6-phosphate 87.0
Fructose 1,6-bisphosphate 22.0
ATP 11,400
ADP 1,320
Source:
From Williamson, J.R. (1965) Glycolytic control mechanisms I:
inhibition of glycolysis by acetate and pyruvate in the isolated, perfused
rat heart.
J. Biol. Chem.
240, 2308–2321.
*Calculated as mmol/mL of intracellular water.
(a) Calculate Q, [fructose 1,6-bisphosphate][ADP]/[fructose 6-phosphate][ATP], for the PFK-1
reaction under physiological conditions.
(b) Given a G for the PFK-1 reaction of 14.2 kJ/mol, calculate the equilibrium constant for this
reaction.
(c) Compare the values of Qand K
eq
. Is the physiological reaction near or far from equilibrium?
Explain. What does this experiment suggest about the role of PFK-1 as a regulatory enzyme?
c15PrinciplesofMetabolicRegulation.qxd 12/6/12 4:35 PM Page S-173

S-174 Chapter 15 Principles of Metabolic Regulation
2. Are All Metabolic Reactions at Equilibrium?
(a) Phosphoenolpyruvate (PEP) is one of the two phosphoryl group donors in the synthesis of ATP
during glycolysis. In human erythrocytes, the steady-state concentration of ATP is 2.24 m
M
, that
of ADP is 0.25 m
M
, and that of pyruvate is 0.051 m
M
. Calculate the concentration of PEP at
25 C, assuming that the pyruvate kinase reaction (see Fig. 13–13) is at equilibrium in the cell.
(b) The physiological concentration of PEP in human erythrocytes is 0.023 m
M
. Compare this with
the value obtained in (a). Explain the significance of this difference.
Answer
c15PrinciplesofMetabolicRegulation.qxd 12/6/12 4:35 PM Page S-174
3. Effect of O
2
Supply on Glycolytic Rates The regulated steps of glycolysis in intact cells can be
identified by studying the catabolism of glucose in whole tissues or organs. For example, the glucose
consumption by heart muscle can be measured by artificially circulating blood through an isolated
intact heart and measuring the concentration of glucose before and after the blood passes through the
heart. If the circulating blood is deoxygenated, heart muscle consumes glucose at a steady rate. When
oxygen is added to the blood, the rate of glucose consumption drops dramatically, then is maintained
at the new, lower rate. Explain.
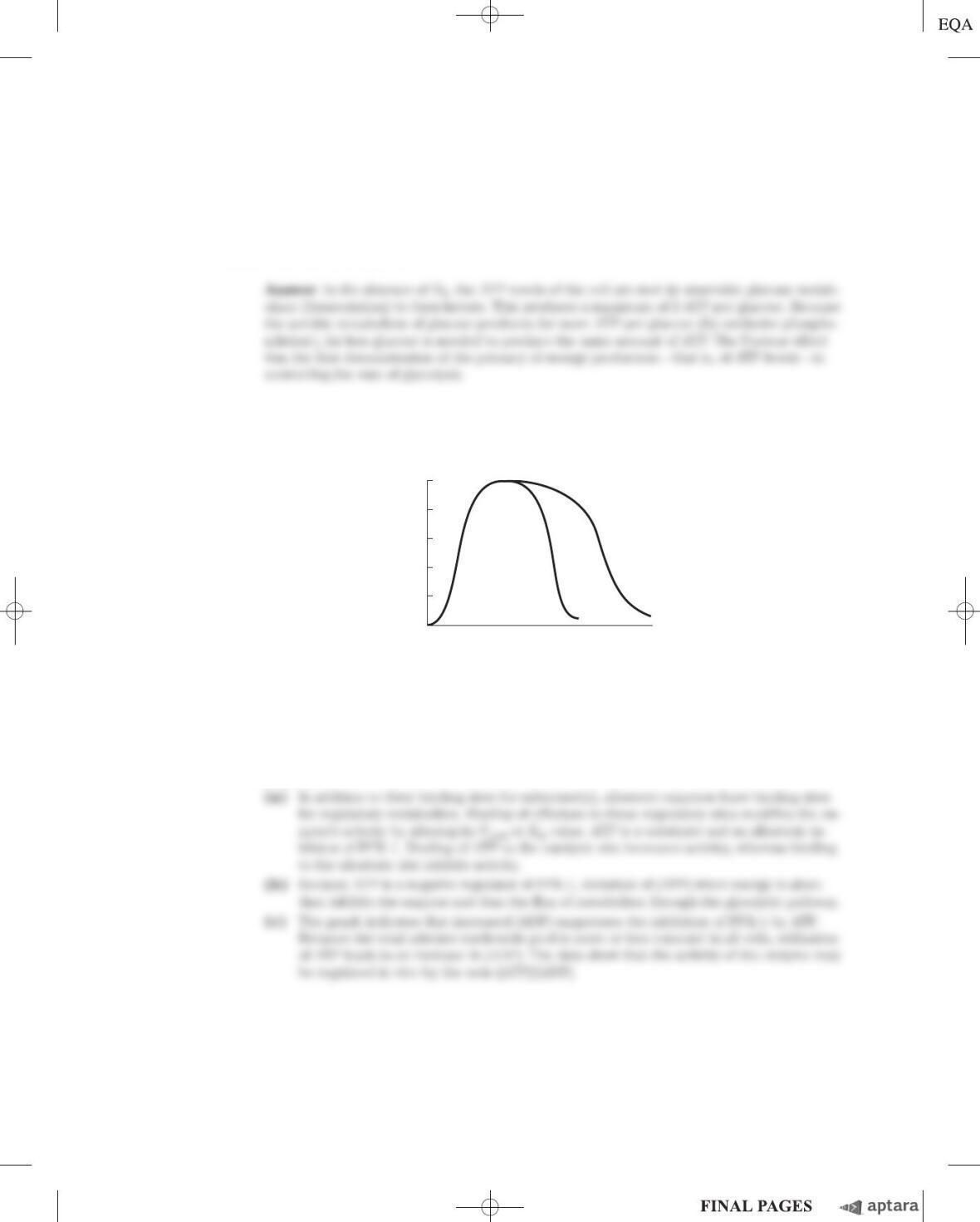
4. Regulation of PFK-1 The effect of ATP on the allosteric enzyme PFK-1 is shown below. For a given
concentration of fructose 6-phosphate, the PFK-1 activity increases with increasing concentrations of
ATP, but a point is reached beyond which increasing the concentration of ATP inhibits the enzyme.
Chapter 15 Principles of Metabolic Regulation S-175
PFK-1 activity (% of Vmax)
[ATP]
High
[ADP]
100
80
60
40
20
0
Low
[ADP]
(a) Explain how ATP can be both a substrate and an inhibitor of PFK-1. How is the enzyme regu-
lated by ATP?
(b) In what ways is glycolysis regulated by ATP levels?
(c) The inhibition of PFK-1 by ATP is diminished when the ADP concentration is high, as shown in
the illustration. How can this observation be explained?
Answer
c15PrinciplesofMetabolicRegulation.qxd 12/6/12 4:35 PM Page S-175

S-176 Chapter 15 Principles of Metabolic Regulation
5. Cellular Glucose Concentration The concentration of glucose in human blood plasma is maintained
at about 5 m
M
. The concentration of free glucose inside a myocyte is much lower. Why is the concentra-
tion so low in the cell? What happens to glucose after entry into the cell? Glucose is administered
intravenously as a food source in certain clinical situations. Given that the transformation of glucose to
glucose 6-phosphate consumes ATP, why not administer intravenous glucose 6-phosphate instead?
Answer Glucose enters cells and is immediately exposed to hexokinase, which converts
6. Enzyme Activity and Physiological Function The V
max
of the enzyme glycogen phosphorylase
from skeletal muscle is much greater than the V
max
of the same enzyme from liver tissue.
(a) What is the physiological function of glycogen phosphorylase in skeletal muscle? In liver tissue?
(b) Why does the V
max
of the muscle enzyme need to be larger than that of the liver enzyme?
Answer
7. Glycogen Phosphorylase Equilibrium Glycogen phosphorylase catalyzes the removal of glucose from
glycogen. The G for this reaction is 3.1 kJ/mol. (a) Calculate the ratio of [P
i
] to [glucose 1-phosphate]
when the reaction is at equilibrium. (Hint: The removal of glucose units from glycogen does not
change the glycogen concentration.) (b) The measured ratio [P
i
]/[glucose 1-phosphate] in myocytes
under physiological conditions is more than 100:1. What does this indicate about the direction of
metabolite flow through the glycogen phosphorylase reaction in muscle? (c) Why are the equilibrium
and physiological ratios different? What is the possible significance of this difference?
Answer

8. Regulation of Glycogen Phosphorylase In muscle tissue, the rate of conversion of glycogen to
glucose 6-phosphate is determined by the ratio of phosphorylase a(active) to phosphorylase b(less
active). Determine what happens to the rate of glycogen breakdown if a muscle preparation containing
glycogen phosphorylase is treated with (a) phosphorylase kinase and ATP; (b) PP1; (c) epinephrine.
Answer
9. Glycogen Breakdown in Rabbit Muscle The intracellular use of glucose and glycogen is tightly
regulated at four points. To compare the regulation of glycolysis when oxygen is plentiful and when it
is depleted, consider the utilization of glucose and glycogen by rabbit leg muscle in two physiological
settings: a resting rabbit, with low ATP demands, and a rabbit that sights its mortal enemy, the coyote,
and dashes into its burrow. For each setting, determine the relative levels (high, intermediate, or low)
of AMP, ATP, citrate, and acetyl-CoA and describe how these levels affect the flow of metabolites
through glycolysis by regulating specific enzymes. In periods of stress, rabbit leg muscle produces
much of its ATP by anaerobic glycolysis (lactate fermentation) and very little by oxidation of acetyl-CoA
derived from fat breakdown.
Chapter 15 Principles of Metabolic Regulation S-177
c15PrinciplesofMetabolicRegulation.qxd 12/6/12 4:35 PM Page S-177

S-178 Chapter 15 Principles of Metabolic Regulation
10. Glycogen Breakdown in Migrating Birds Unlike the rabbit with its short dash, migratory birds
require energy for extended periods of time. For example, ducks generally fly several thousand miles
during their annual migration. The flight muscles of migratory birds have a high oxidative capacity and
obtain the necessary ATP through the oxidation of acetyl-CoA (obtained from fats) via the citric acid
cycle. Compare the regulation of muscle glycolysis during short-term intense activity, as in the fleeing
rabbit, and during extended activity, as in the migrating duck. Why must the regulation in these two
settings be different?
Answer Migratory birds have a very efficient respiratory system to ensure that O
2
is available
11. Enzyme Defects in Carbohydrate Metabolism Summaries of four clinical case studies follow. For
each case, determine which enzyme is defective and designate the appropriate treatment, from the
lists provided at the end of the problem. Justify your choices. Answer the questions contained in each
case study. (You may need to refer to information in Chapter 14.)
Case A The patient develops vomiting and diarrhea shortly after milk ingestion. A lactose tolerance
test is administered. (The patient ingests a standard amount of lactose, and the glucose and galactose
concentrations of blood plasma are measured at intervals. In individuals with normal carbohydrate me-
tabolism, the levels increase to a maximum in about 1 hour, then decline.) The patient’s blood glucose
and galactose concentrations do not increase during the test. Why do blood glucose and galactose in-
crease and then decrease during the test in healthy individuals? Why do they fail to rise in the patient?
Case B The patient develops vomiting and diarrhea after ingestion of milk. His blood is found to
have a low concentration of glucose but a much higher than normal concentration of reducing sugars.
The urine tests positive for galactose. Why is the concentration of reducing sugar in the blood high?
Why does galactose appear in the urine?
Case C The patient complains of painful muscle cramps when performing strenuous physical
exercise but has no other symptoms. A muscle biopsy indicates a muscle glycogen concentration much
higher than normal. Why does glycogen accumulate?
Case D The patient is lethargic, her liver is enlarged, and a biopsy of the liver shows large
amounts of excess glycogen. She also has a lower than normal blood glucose level. What is the reason
for the low blood glucose in this patient?
Defective Enzyme
(a) Muscle PFK-1
(b) Phosphomannose isomerase
(c) Galactose 1-phosphate uridylyltransferase
(d) Liver glycogen phosphorylase

(e) Triose kinase
(f) Lactase in intestinal mucosa
(g) Maltase in intestinal mucosa
(h) Muscle debranching enzyme
Treatment
1. Jogging 5 km each day
2. Fat-free diet
3. Low-lactose diet
4. Avoiding strenuous exercise
5. Large doses of niacin (the precursor of NAD
)
6. Frequent feedings (smaller portions) of a normal diet
12. Effects of Insufficient Insulin in a Person with Diabetes A man with insulin-dependent diabetes
is brought to the emergency room in a near-comatose state. While vacationing in an isolated place, he
lost his insulin medication and has not taken any insulin for two days.
(a) For each tissue listed below, is each pathway faster, slower, or unchanged in this patient,
compared with the normal level when he is getting appropriate amounts of insulin?
(b) For each pathway, describe at least one control mechanism responsible for the change you predict.
Tissue and Pathways
1. Adipose: fatty acid synthesis
2. Muscle: glycolysis; fatty acid synthesis; glycogen synthesis
3. Liver: glycolysis; gluconeogenesis; glycogen synthesis; fatty acid synthesis; pentose phosphate
pathway
Chapter 15 Principles of Metabolic Regulation S-179
c15PrinciplesofMetabolicRegulation.qxd 12/6/12 4:35 PM Page S-179

S-180 Chapter 15 Principles of Metabolic Regulation
13. Blood Metabolites in Insulin Insufficiency For the patient described in Problem 12, predict the
levels of the following metabolites in his blood before treatment in the emergency room, relative to lev-
els maintained during adequate insulin treatment: (a) glucose; (b) ketone bodies; (c) free fatty acids.
14. Metabolic Effects of Mutant Enzymes Predict and explain the effect on glycogen metabolism of
each of the following defects caused by mutation: (a) loss of the cAMP-binding site on the regulatory
subunit of protein kinase A (PKA); (b) loss of the protein phosphatase inhibitor (inhibitor 1 in Fig. 15–42);
(c) overexpression of phosphorylase bkinase in liver; (d) defective glucagon receptors in liver.
Answer
15. Hormonal Control of Metabolic Fuel Between your evening meal and breakfast, your blood glucose
drops and your liver becomes a net producer rather than consumer of glucose. Describe the hormonal
basis for this switch, and explain how the hormonal change triggers glucose production by the liver.
16. Altered Metabolism in Genetically Manipulated Mice Researchers can manipulate the genes of a
mouse so that a single gene in a single tissue either produces an inactive protein (a “knockout” mouse)
or produces a protein that is always (constitutively) active. What effects on metabolism would you pre-
dict for mice with the following genetic changes: (a) knockout of glycogen debranching enzyme in the
liver; (b) knockout of hexokinase IV in liver; (c) knockout of FBPase-2 in liver; (d) constitutively active
FBPase-2 in liver; (e) constitutively active AMPK in muscle; (f) constitutively active ChREBP in liver?

Answer
Data Analysis Problem
17. Optimal Glycogen Structure Muscle cells need rapid access to large amounts of glucose during heavy
exercise. This glucose is stored in liver and skeletal muscle in polymeric form as particles of glycogen.
The typical glycogen particle contains about 55,000 glucose residues (see Fig. 15–35b). Meléndez-Hevia,
Waddell, and Shelton (1993) explored some theoretical aspects of the structure of glycogen, as de-
scribed in this problem.
(a) The cellular concentration of glycogen in liver is about 0.01
M
. What cellular concentration of
free glucose would be required to store an equivalent amount of glucose? Why would this con-
centration of free glucose present a problem for the cell?
Glucose is released from glycogen by glycogen phosphorylase, an enzyme that can remove glucose
molecules, one at a time, from one end of a glycogen chain. Glycogen chains are branched (see
Figs 15–28 and 15–35b), and the degree of branching—the number of branches per chain—has a
powerful influence on the rate at which glycogen phosphorylase can release glucose.
(b) Why would a degree of branching that was too low (i.e., below an optimum level) reduce the rate
of glucose release? (Hint: Consider the extreme case of no branches in a chain of 55,000 glucose
residues.)
(c) Why would a degree of branching that was too high also reduce the rate of glucose release?
(Hint: Think of the physical constraints.)
Meléndez-Hevia and colleagues did a series of calculations and found that two branches per chain
(see Fig. 15–35b) was optimal for the constraints described above. This is what is found in glycogen
stored in muscle and liver.
To determine the optimum number of glucose residues per chain, Meléndez-Hevia and coauthors
considered two key parameters that define the structure of a glycogen particle: tthe number of tiers
of glucose chains in a particle (the molecule in Fig. 15–35b has five tiers); g
c
the number of glucose
residues in each chain. They set out to find the values of tand g
c
that would maximize three quantities:
(1) the amount of glucose stored in the particle (G
T
) per unit volume; (2) the number of unbranched
glucose chains (C
A
) per unit volume (i.e., number of chains in the outermost tier, readily accessible to
glycogen phosphorylase); and (3) the amount of glucose available to phosphorylase in these unbranched
chains (G
PT
).
(d) Show that C
A
2
t1
. This is the number of chains available to glycogen phosphorylase before
the action of the debranching enzyme.
(e) Show that C
T
, the total number of chains in the particle, is given by C
T
2
t
1. Thus G
T
g
c
(C
T
) g
c
(2
t
1), the total number of glucose residues in the particle.
(f) Glycogen phosphorylase cannot remove glucose from glycogen chains that are shorter than five
glucose residues. Show that G
PT
(g
c
4)(2
t1
). This is the amount of glucose readily avail-
able to glycogen phosphorylase.
(g) Based on the size of a glucose residue and the location of branches, the thickness of one tier of
glycogen is 0.12g
c
nm 0.35 nm. Show that the volume of a particle, V
s
, is given by the
equation V
s
4
3
t
3
(0.12g
c
0.35)
3
nm
3
.
Chapter 15 Principles of Metabolic Regulation S-181
c15PrinciplesofMetabolicRegulation.qxd 12/6/12 4:35 PM Page S-181

S-182 Chapter 15 Principles of Metabolic Regulation
Meléndez-Hevia and coauthors then determined the optimum values of tand g
c
—those that gave
the maximum value of a quality function, f, that maximizes G
T
, C
A
, and G
PT
, while minimizing
V
S
: f
G
T
C
V
A
s
G
PT
. They found that the optimum value of g
c
is independent of t.
(h) Choose a value of tbetween 5 and 15 and find the optimum value of g
c
. How does this compare
with the g
c
found in liver glycogen (see Fig. 15–35b)? (Hint: You may find it useful to use a
spreadsheet program.)
Answer